DNA origami
Distinguish between functional dynamics of DNA origami and misfolded defects in real time
Characterise DNA origami nanostructures with single-molecule precision
DNA origami nanostructures have a wide range of applications, from biosensing to drug delivery and the formation of synthetic nanopores. Their structure is carefully designed to ensure that DNA scaffolds fold into the correct shape and are responsive to intended stimuli. Common techniques used to characterise DNA origami nanostructures include atomic force microscopy, Cryo-EM and gel electrophoresis – methods that produce static and potentially artefactual structures or result in ensemble averages, potentially missing crucial intermediates and conformational dynamics.
By tracking single molecules in solution, researchers can map dynamic conformational shifts, defining salt or pH responsiveness while identifying misfolded nanostructures, even those at low abundance.
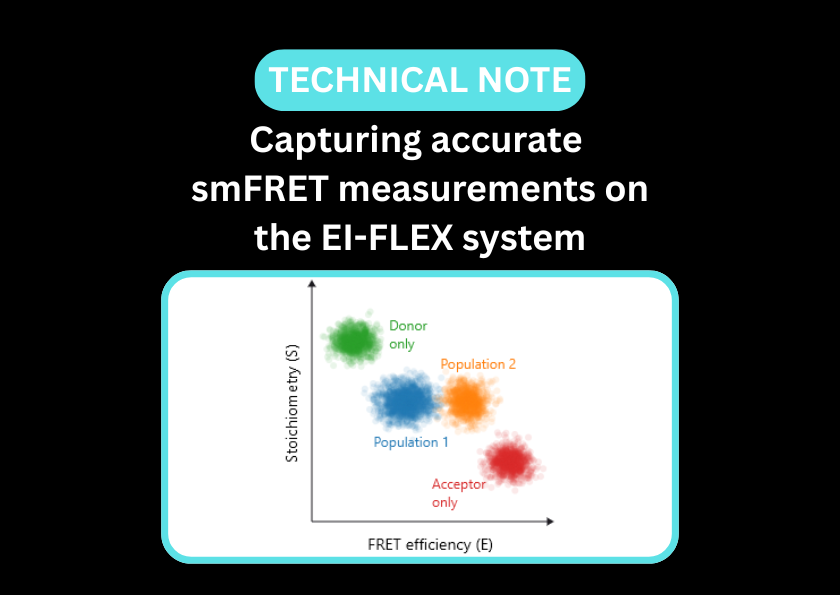
Single-molecule Förster resonance energy transfer (smFRET) serves as a critical tool for the characterisation of DNA nanostructures. Beyond performing quality checks to identify structural heterogeneity, smFRET is used to observe the active operation of DNA-based devices, including the locomotion of DNA walkers, the actuation of DNA tweezers, and the responsiveness of nanovehicles and biosensors.
Additionally, Fluorescence Correlation Spectroscopy (FCS) is utilised to quantify the efficiency of the self-assembly process, distinguishing between fully formed scaffolds, incomplete intermediates, and monomeric staple/scaffold components by analysing their co-diffusion and brightness profiles.
Validate the structural integrity and dynamic functionality of DNA origami scaffolds
- Measure DNA nanostructures in solution without tethering - be certain the function of your nanostructures isn’t a result of surface-induced artefacts
- Obtain structural data alongside stoichiometry and concentration, all on the same platform
- Verify design accuracy to ensure that the folded origami structure matches the in silico model
- Monitor transition rates between open and closed states in response to external stimuli (e.g., DNA fuel strands, pH, or ions)
- Directly measure the concentration of correctly assembled origami structures versus excess staples in solution
- Characterise hierarchical self-assembly and quantify payload incorporation
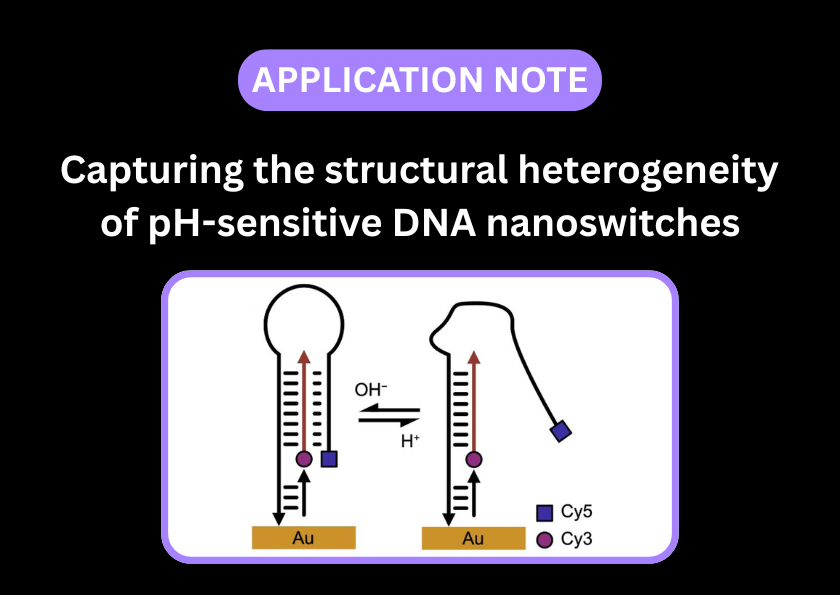
smFRET can characterise the effect of pH on the opening and closing of a DNA nanoswitch
smFRET is ideal for quantifying the functionality of DNA nanoswitches; by analysing single molecules, heterogeneous populations of open and closed nanoswitches can be distinguished from those that are not folding correctly.
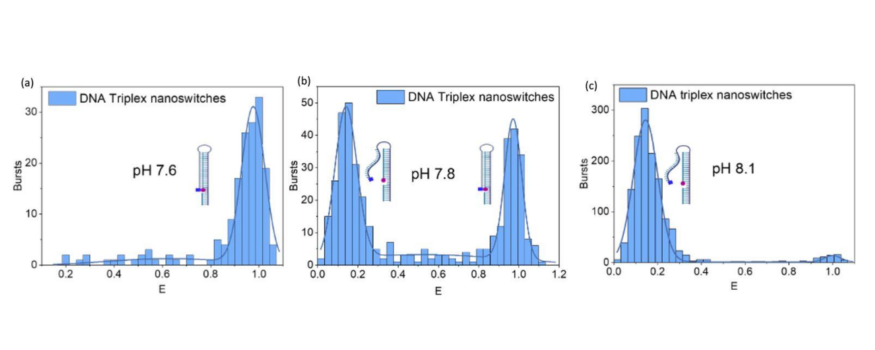
Figure 1 – smFRET reveals heterogeneity in pH-responsive DNA nanoswitches
a) At pH 7.6, the DNA nanoswitch is closed (high-FRET population)
b) At pH 7.8, there is a heterogeneous population of open and closed nanoswitches (low and high-FRET populations)
c) At pH 8.1, all DNA nanoswitches are open (low-FRET p`opulation)
Figure taken from: D’Rozario, F. et al. Electronic Actuation of Surface-Immobilized, pH-Responsive DNA Nanoswitches. ACS Appl. Mater. Interfaces 18, 18039–18048 (2026)
DNA origami papers and pre-prints featuring EI-FLEX data
DNA origami FAQs
Can smFRET monitor the dynamic reconfiguration of DNA origami?
Many origami structures are designed to be dynamic. By placing a FRET pair across the moving interface, you can watch the structure switch between open and closed states in response to external triggers like pH, ions, or other DNA strands.
How does FCS measure the assembly and purity of DNA origami?
FCS is used to check if the scaffold and staples have successfully formed a single, compact structure. An assembled origami diffuses more slowly than individual DNA staples. By measuring the diffusion time and the molecular brightness, you can distinguish between a fully formed nanostructure and a failed assembly of loose strands.
Can FCCS confirm the incorporation of proteins or nanoparticles onto origami?
By labelling the DNA origami and the protein cargo with separate fluorophores, FCCS can detect the co-diffusion of a protein with the DNA scaffold when successfully docked. This is faster and more accurate than traditional gel mobility shift assays.
Can I use smFRET to study the breathing or flexibility of an origami structure?
Yes – by analysing the variance of FRET efficiencies (using burst variance analysis), you can determine the mechanical stiffness of the nanostructure and how it changes in different buffers.
Can I detect the opening of DNA origami drug-delivery nanostructures?
smFRET can distinguish between the open and closed states of nanostructures. When a trigger (like a specific mRNA sequence or a protein) binds the nanostructure and causes it to release its cargo, this conformational change can be captured via shifts in FRET efficiencies. FCCS would also reveal a loss in codiffusion of fluorescent signal if both the DNA origami and its cargo were labelled.
Is FCS sensitive enough to measure the hydrodynamic radius of different origami shapes?
Yes, FCS can detect changes in hydrodynamic radius, which depend on both molecular size and shape. Distinguishing between different shapes of DNA origami of similar size is feasible.
Can I use these tools to study hierarchical assembly?
FCCS can be used to monitor the assembly order of two differentially labelled origami tiles and build accurate models of hierarchical assembly.
How does photobleaching affect origami studies in smFRET?
All fluorophores are susceptible to undesired photophysical effects such as photobleaching. This can be mitigated by adding oxygen scavenging systems to buffers or by limiting the extent of laser excitation. The small excitation volume and minimal duration of excitation during confocal smFRET experiments limit the effects of photobleaching, reducing the dependence on oxygen scavenging systems that may cause undesirable changes in buffer conditions.
How is smFRET used to characterise the function of DNA tweezers?
smFRET can monitor the structural transitions of DNA tweezers by attaching fluorophores to the device’s arms. Unlike static imaging, smFRET can track the open and closed states in real-time, providing quantitative data on the kinetics of actuation and the efficiency of the switching mechanism. This enables the identification of intermediate states and structural heterogeneity that might be missed by ensemble measurements.
Can confocal smFRET complement AFM or cryo-EM for studying DNA origami?
While AFM and cryo-EM provide high-resolution static DNA origami structures, confocal smFRET captures the DNA origami’s behaviour in physiologically-relevant conditions, capturing fast transitions that are often missed by other structural techniques.
"It has opened up new avenues of research because of the combined FCS, FCCS and single-molecule FRET capabilities. The high performance, stability and precision of the EI-FLEX instrument have really been a game-changer for experiments where sensitivity to picomolar concentration of biomolecules really matters."
Dr. Steven Quinn, University of York

Other resources you might be interested in