Intrinsically disordered proteins
Decoding the architecture of disorder in the dark proteome
Mapping the conformational landscape: From random coils to pathological aggregates
The study of intrinsically disordered proteins (IDPs) presents a fundamental paradox in structural biology: how do we define the function of a protein that has no fixed structure? Unlike globular proteins, IDPs exist as highly dynamic ensembles, constantly fluctuating between expanded random coil states and transiently collapsed conformations. This inherent plasticity makes them notoriously difficult to characterise. To truly understand the structure of an IDP, it is crucial to map the entire conformational landscape.
Single-molecule techniques provide the high-resolution data necessary to decode this area of the dark proteome by isolating individual molecular behaviours from the noise of the bulk population. Specifically, single-molecule Förster resonance energy transfer (smFRET) acts as a sensitive molecular ruler, measuring the distance between specific residues to reveal the full distribution of states within a conformational ensemble.
Fluorescence correlation spectroscopy (FCS) and fluorescence cross-correlation spectroscopy (FCCS) complement this by quantifying the global expansion or compaction of the protein chain as it responds to changes in the cellular environment or post-translational modifications, while also characterising the IDP interactome. Together, this structural and binding information is vital for understanding IDP function and how this disorder contributes to disease.
Characterise the complex conformational landscape and interactome of IDPs at single-molecule precision
- Measure the hydrodynamic volume of an IDP with high precision to determine how the protein behaves under different environmental conditions
- Resolve conformational ensembles using smFRET to reveal whether an IDP is a single broad population or if it fluctuates between distinct sub-states
- Map segmental flexibility to identify specific disordered regions that are highly dynamic and others that might harbour transient structural elements
- Observe the transition from a disordered state to a structured fold upon ligand or chaperone binding
- Characterise fuzzy complex stoichiometry, understanding how IDPs bind multiple partners simultaneously or form transient, non-specific complexes
- Detect early-stage compaction and misfolding, serving as an early warning system for the formation of toxic oligomers
- Monitor material transitions in phase separation; FCS can be used to measure the change in molecular mobility as the protein moves from the dilute phase into a dense droplet
smFRET supports the mapping of IDP ensemble structures
Naudi-Fabra et al. combined smFRET, FRET-lifetimes, NMR (chemical shifts and PREs), and SAXS to quantitatively map the conformational ensemble of the disordered domain of the measles virus phosphoprotein. smFRET provided highly complementary data, unifying residue-specific NMR data and global SAXS dimensions to produce a dynamic conformational ensemble.
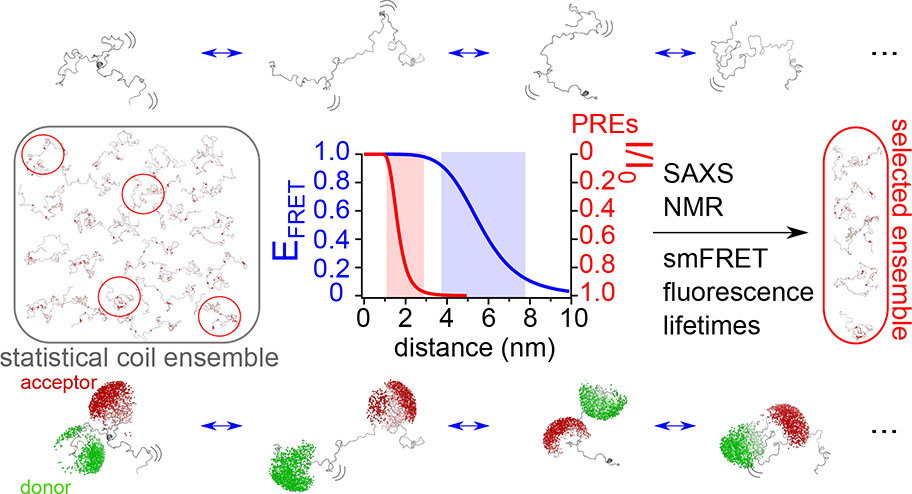
By combining multiple structural and biophysical techniques with smFRET, a predicted set of conformations undertaken by the IDP measles virus phosphoprotein could be derived.
Figure taken from Naudi-Fabra et al., 2021
Intrinsically disordered protein resources


Intrinsically disordered protein FAQs
How does smFRET complement NMR to study IDP conformations?
While NMR provides excellent local residue-specific information, it yields an ensemble average that can mask rare but biologically significant compact or expanded states. smFRET captures inter-dye distances within individual molecules, allowing users to see if an IDP exists as a single broad conformational ensemble or multiple distinct populations.
Can we observe the transition from disorder to order upon binding?
Yes – smFRET can monitor an IDP as it binds to a partner. If the binding partner induces a more stable confirmation in the IDP, smFRET data will show a less dynamic signal, alongside a shift in FRET efficiency.
Can single-molecule techniques distinguish between induced fit and conformational selection in IDPs?
By analysing the timescales of conformational fluctuations with smFRET and FCS in the presence of varying ligand concentrations, we can determine if the protein folds after binding (induced fit) or if the partner selects a pre-existing folded state from the IDP’s natural ensemble.
How does FCCS identify IDP-binding interactions?
IDPs often bind to partners without fully folding. FCCS can quantify the binding affinity of these interactions in solution, even when the interaction is transient or low-affinity, by monitoring the co-diffusion of the IDP and its partner.
Can this technology study liquid-liquid phase separation (LLPS) in lysates?
Yes. While LLPS is a cellular process, it is most rigorously studied in vitro using purified proteins or cell lysates. FCS can measure the internal viscosity of these droplets by tracking how fast molecules move inside them, identifying the moment a liquid droplet transitions into a rigid, gel-like state.
Can I study the effect of post-translational modifications (PTMs) on IDP structure?
PTMs, such as phosphorylation, often act as molecular switches for IDPs. Single-molecule analysis can show how a single phosphate group shifts the conformational landscape; for example, by triggering a global expansion of the protein chain that prevents aggregation. The extent of phosphorylation, or other PTMs, can also be quantified.
How do IDPs behave in molecular crowding conditions?
The cellular interior is packed with macromolecules that can force an IDP to collapse. We can simulate this by adding crowders (like PEG or Ficoll) and using smFRET to measure the change in the IDP’s dimensions or FCS to quantify the hydrodynamic radius, providing a more biologically relevant picture than dilute buffer studies.
What should be considered when labelling IDPs with fluorophores?
Since IDPs lack a hydrophobic core, they are sensitive to labelling. It is recommended to use small, hydrophilic fluorophores and site-specific labelling (e.g., unnatural amino acids or maleimide chemistry) at positions that are bioinformatically predicted to have minimal impact on the protein’s native disorder.
Can I study IDPs that are only unfolded under certain conditions (e.g., pH or salts)?
Yes – you can perform single-molecule titrations, watching the protein’s conformational ensemble shift in real-time as you vary factors such as pH or salt concentration to find the exact midpoint of a folding/unfolding transition.
How does smFRET complement small-angle X-ray scattering (SAXS) when characterising IDPs?
While SAXS is an excellent tool for determining the global envelope and radius of gyration of a protein, it provides an ensemble average that can be difficult to interpret for highly flexible IDPs. smFRET acts as a powerful internal validation tool by providing precise, site-specific distance distributions between defined residues.
"It has opened up new avenues of research because of the combined FCS, FCCS and single-molecule FRET capabilities. The high performance, stability and precision of the EI-FLEX instrument have really been a game-changer for experiments where sensitivity to picomolar concentration of biomolecules really matters."
Dr Steven Quinn, University of York

Other resources you might be interested in


