Membrane Proteins
Capture membrane proteins in motion at single-molecule sensitivity
Navigating the challenges of membrane proteins
Membrane proteins are the vital conduits of cellular life, serving as receptors, transporters, and ion channels that regulate communication between the cell and its environment. While they represent over 60% of current drug targets, notably GPCRs, they remain notoriously difficult to study as they require a lipid environment to maintain structural integrity.
Traditional structural biology methods often struggle with membrane proteins because they are prone to instability when extracted from the bilayer, and their large, complex transmembrane domains make them resistant to crystallisation. Furthermore, because these proteins function through subtle, high-speed conformational shifts, static snapshots often fail to capture the full mechanism of how they move, bind, and signal.
Single-molecule spectroscopy offers a transformative solution by allowing researchers to observe membrane proteins within native-like environments, such as detergent micelles, nanodiscs, or lipid vesicles. Single-molecule Förster resonance energy transfer (smFRET) acts as a dynamic molecular ruler, capturing the real-time structural transitions of receptors as they bind ligands or transporters as they shuttle cargo across a membrane. Meanwhile, fluorescence correlation spectroscopy (FCS) offers critical insights into the protein’s behaviour in a fluid, living bilayer, such as whether a receptor functions as a monomer or a dimer, or how its diffusion changes in response to drug binding.
Analyse membrane proteins in nanodiscs and vesicles with single-molecule spectroscopy
- Map conformational shifts, measuring the distance between extracellular and intracellular domains in real-time
- Measure ligand-binding affinity in lipid environments
- Quantify oligomerisation states of receptors, such as GPCRs, that function as dynamic monomers, dimers, or higher-order oligomers
- Resolve rare intermediate states, such as short-lived intermediate states that occur during the opening or closing of an ion channel
- Characterise protein-lipid interactions, such as the allosteric modulation of membrane proteins by lipids
- Track lateral mobility and membrane fluidity, measuring the diffusion speed of a protein as it moves through a bilayer
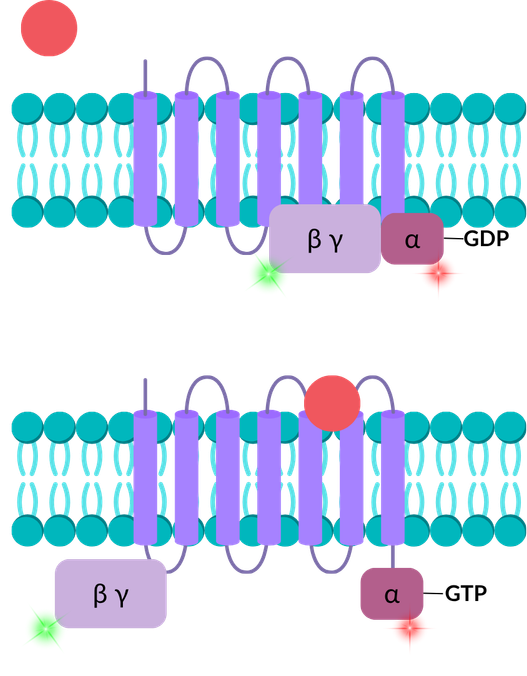
Understand ligand-induced conformational changes in GPCRs
smFRET, FCS and FCCS can support assays investigating the impact of ligand binding to GPCRs, such as agonists and antagonists. In this illustrative example, green and red dyes have been placed on the α and β/γ subunits of a GPCR, respectively, permitting analysis by smFRET. Prior to ligand binding, the subunits are in a complex, producing a high-FRET signal. This complex is disrupted by the binding of a ligand, which is captured as a low-FRET signal. Ligand affinity can also be measured by FCS if the ligand is labelled with a dye.
Membrane protein papers and pre-prints featuring EI-FLEX data


Membrane protein FAQs
Can I study membrane proteins in their native lipid environment?
Yes – our technology allows you to study proteins in nanodiscs, liposomes, or detergent micelles. By using these lipid-mimetic systems, you can ensure the protein maintains its native fold and biological activity during measurement.
How do single-molecule methods handle the high background of lipid vesicles?
Because this technology uses a highly focused confocal volume (less than one femtoliter), only fluorescently labelled molecules that are diffusing through this volume are analysed, filtering out high background interference. This permits the acquisition of data in complex biofluids, such as cell lysates or nanodiscs.
Is it possible to study how membrane composition (e.g., cholesterol) affects protein function?
This is a major strength of the platform. You can systematically vary the lipid ratios in your nanodiscs or vesicles and use smFRET to watch how specific lipids trigger conformational changes in a receptor, or use FCS to see how membrane fluidity alters the protein’s lateral mobility.
Can I distinguish between active and inactive states of a GPCR?
Yes. By placing a FRET pair on the intracellular loops or the C-terminus, you can monitor the distance changes that occur upon ligand binding. This distinguishes between full agonists, partial agonists, and antagonists based on the specific conformational fingerprint they induce in the receptor.
How can I determine if a receptor is a monomer or a dimer?
FCS and Photon Counting Histogram (PCH) analysis are used to measure the molecular brightness of the particles. A dimer will effectively have twice the brightness of a monomer. Additionally, FCCS can be used with differently coloured labels to see if two protein subunits are physically moving together as a stable complex.
Can this technology resolve the allosteric effects of drug candidates?
Yes. Because smFRET analyses individual molecules, it can detect subtle structural shifts caused by allosteric modulators that might be missed in bulk assays. This is particularly useful for identifying drugs that don’t compete with the primary ligand but change the protein’s dwell time in a specific active state.
Can I observe a transporter flip between inward- and outward-facing states?
This is a classic application for smFRET. By labelling the gates of a transporter, you can watch the real-time transition between different structural states. This provides a dynamic view of the transport cycle, supporting static structural data.
Is the system sensitive enough to detect rare, short-lived intermediate states?
Single-molecule detection permits the analysis of heterogeneous populations, identifying even rare, transient conformations or those that are dynamically interconverting – all within the same sample.
Can I measure the binding of small molecules to a transporter?
FCS is ideal for quantifying the binding affinity of labelled ligands to the transporter. Additionally, FCCS can be used to cross-correlate the signal of a fluorescently labelled ligand with the labelled membrane protein to understand the stoichiometry of this interaction.
How much protein is required for a single-molecule membrane study?
Very little, which is a huge advantage since membrane proteins are so difficult to express and purify. A typical measurement requires only a few microliters of sample at picomolar to nanomolar concentrations.
How does the system handle sticky membrane proteins that aggregate easily?
FCS is ideal for identifying aggregating proteins. It will show if a protein is monodisperse (one clean peak) or if it has started to aggregate (bright, slow-moving spikes in the data), allowing you to optimise your buffer or detergent conditions in real-time.
“Solution-based measurements, like those taken using the EI-FLEX, avoid surface tethering, which can introduce artefacts, especially for proteins that don’t behave well near hydrophobic surfaces. Keeping proteins in solution lets you observe their native conformation, and it’s much easier experimentally: you label, dilute, and measure, often getting results in minutes rather than days or weeks with TIRF-based systems."
Associate Professor Kuang Shen, UMass Chan

Other resources you might be interested in


