For decades, biophysics and structural biology operated under a strict constraint: if you wanted to see a molecule clearly, you had to freeze it, crystallise it, or look at billions of them simultaneously. Biochemistry was built on ensemble averages, a methodology that treats molecular populations as uniform entities.
But biology in the cell is chaotic, dynamic and remarkably individualised.
The development of single-molecule Förster Resonance Energy Transfer (smFRET) empowered the field to move beyond ensemble data. By tracking the non-radiative energy transfer between donor and acceptor fluorophores on individual molecules, smFRET transformed how we investigate the subcellular world. It changed our perspective from static snapshots to real-time, high-resolution observations of molecular mechanisms in action.
As we celebrate the 30th anniversary of its first publication, this retrospective article traces the chronological roadmap of smFRET. We examine the seminal papers, a series of technological advances, and the foundational hardware and software that evolved a niche biophysical tool into an indispensable, and now accessible, engine of modern molecular discovery.
30 years at a glance
(Pre-smFRET) 1967 ─ Stryer & Haugland define the bulk ‘spectroscopic ruler’
(Pre-smFRET) 1976 – Hirschfeld optical tracking breaks the single-molecule barrier
1996 – Ha et al. realise single-molecule FRET on surface-tethered DNA
1999 – The technique expands to dynamic biological systems (RNA folding & enzyme cycles)
2000 – Deniz et al. introduce free-diffusion solution burst analysis
2004 – Kapanidis et al. introduce ALEX; Hohng et al. expand coordinates via 3-colour FRET
2006 – ‘DNA Scrunching’ mechanical model definitively proven during transcription initiation
2012 – Community software (SMART, FRETBursts) unifies open data analysis standards
2015 – In vivo dynamics and full mammalian GPCR signalling captured in active cell matrices
2020 – The ‘smfBox’ provides open-source hardware for smFRET
2021+ ─ Exciting Instruments commercialises the EI-FLEX and EI-FLEX Pro, introducing automated, ambient-light benchtop single-molecule screening to life sciences and drug discovery
Precursors to smFRET (1967–1976)
Before smFRET could come to be, two foundational pillars had to converge: the quantitative physics of energy transfer and the optical capacity to detect a single photon emitted from an isolated small molecule.
The spectroscopic ruler
In 1967, Lubert Stryer and Richard Haugland published their landmark study, ‘Energy transfer: a spectroscopic ruler’1. Working in bulk solutions, they attached energy donor and acceptor groups to prolines of varying oligomer lengths.
Their findings validated Theodor Förster’s long-held theory: the efficiency of energy transfer (E) drops off inversely with the sixth power of the distance (R) separating the dyes.
Here, R0 represents the Förster distance at which the transfer efficiency is exactly 50%.

Stryer and Haugland coined the term ‘spectroscopic ruler’, establishing a quantitative framework that allowed researchers to infer Ångstrom-level distance changes based on shifting fluorescence signals. Yet, for nearly thirty years, this elegant ruler remained bound to ensemble systems, measuring the statistical average rather than individual molecules.
Isolating the single molecule
The physical capability to see an individual molecule with light arrived a decade later. In 1976, Tomas Hirschfeld published ‘Optical microscopic observation of single small molecules’2. By addressing signal-to-noise limitations through spatial discrimination and high-numerical-aperture optics, Hirschfeld proved that individual fluorophores could yield detectable signals above background scattering.
This paper provided the ultimate technical proof-of-concept that the biophysics community needed: the single molecule was no longer just a statistical abstraction, but rather a tangible, observable physical reality.
The genesis of smFRET (1996–1999)
In 1996, the theoretical concept of the single-molecule spectroscopic ruler became an experimental reality.
The seminal breakthrough
Working in the laboratory of Shuming Chu and Shimon Weiss, Taekjip Ha and his co-workers published the breakthrough paper, ‘Probing the interaction between two single molecules — fluorescence resonance energy transfer between a single donor and a single acceptor’3.
The team immobilised dual-labelled DNA molecules onto a glass surface and analysed them with near-field scanning optical microscopy. By exciting a single donor dye and monitoring the anti-correlated emission of both donor and acceptor channels over time, they mapped the first real-time trajectories of single-pair FRET, proving that the distance ruler could successfully operate at the single-molecule level.
Moving beyond rigid DNA
The immediate challenge after 1996 was proving that smFRET could do more than track distance along a rigid, predictable double-helix DNA standard. The field needed to show that it could resolve actual biological behaviour.
In 1999, Ha et al. achieved this milestone with their paper ‘Ligand-induced conformational changes observed in single RNA molecules’4. This work tracked individual three-helix RNA junctions undergoing conformational folding and unfolding. By monitoring structural shifts induced by magnesium or ribosomal protein binding, the authors proved that tracking the motion of a single molecule undergoing functional conformational transformations was completely achievable.
Concurrently, the technique migrated from nucleic acids to proteins. Ha, et al. published ‘Single-molecule fluorescence spectroscopy of enzyme conformational dynamics and cleavage mechanism’5. The team used intramolecular smFRET to map the resting conformational dynamics of staphylococcal nuclease, alongside intermolecular smFRET to directly witness the binding of donor-labelled enzymes to acceptor-labelled DNA substrates.
This work verified that smFRET could characterise the fast equilibrium fluctuations and catalytic cycles of complex enzymes, cementing its utility for general protein biochemistry.
From surfaces to in-solution expansion of coordinates (2000–2006)
As the field entered the 2000s, two primary challenges emerged. First, surface immobilisation risked altering native biomolecular conformations via non-specific glass interactions. Second, it was difficult to determine whether data was a result of true biomolecular distance changes, or whether incomplete labelling was interfering with the results.
Solution-based confocal dynamics
In 2000, Deniz et al. bypassed surface artefacts entirely with ‘Single-molecule protein folding: Diffusion fluorescence resonance energy transfer studies of the denaturation of chymotrypsin inhibitor 2’6. By utilising solution-based confocal spectroscopy, molecules were allowed to freely diffuse through a femtolitre-sized optical detection volume. Each passing molecule yielded a rapid burst of photons. Free from any surface perturbation, this technique allowed accurate profiling of sub-population distributions during protein denaturation.
ALEX and multi-colour coordinates
In 2004, the capability to analyse heterogeneous samples in solution took a massive technological leap forward. Achillefs Kapanidis, Shimon Weiss, and their colleagues introduced Alternating-Laser Excitation in their paper, ‘Fluorescence-aided molecule sorting: Analysis of structure and interactions by alternating-laser excitation of single molecules’7. Standard smFRET cannot easily distinguish a low-FRET molecule from a molecule missing a functional acceptor dye. ALEX solved this by rapidly interleaving two distinct lasers to directly excite both the donor and the acceptor sequentially during a single diffusion burst. This produced a two-dimensional plot mapping both FRET efficiency (E) and dye stoichiometry (S).
Researchers could instantly filter out incompletely labelled species, so that only data from dual-labelled complexes could be analysed.
The first E/S plot shows all bursts, while the second plot shows those with an S value of ~0.5, indicating doubly labelled molecules.
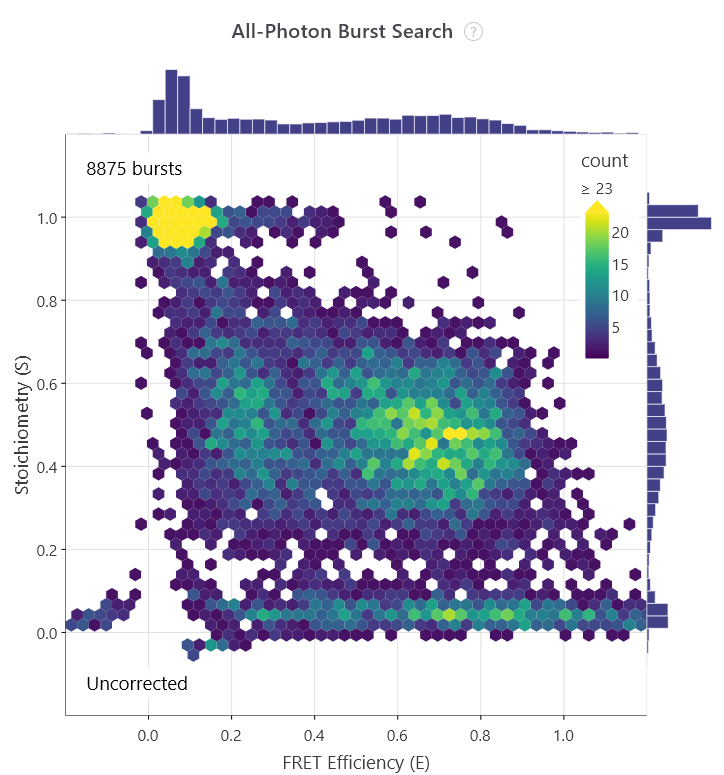
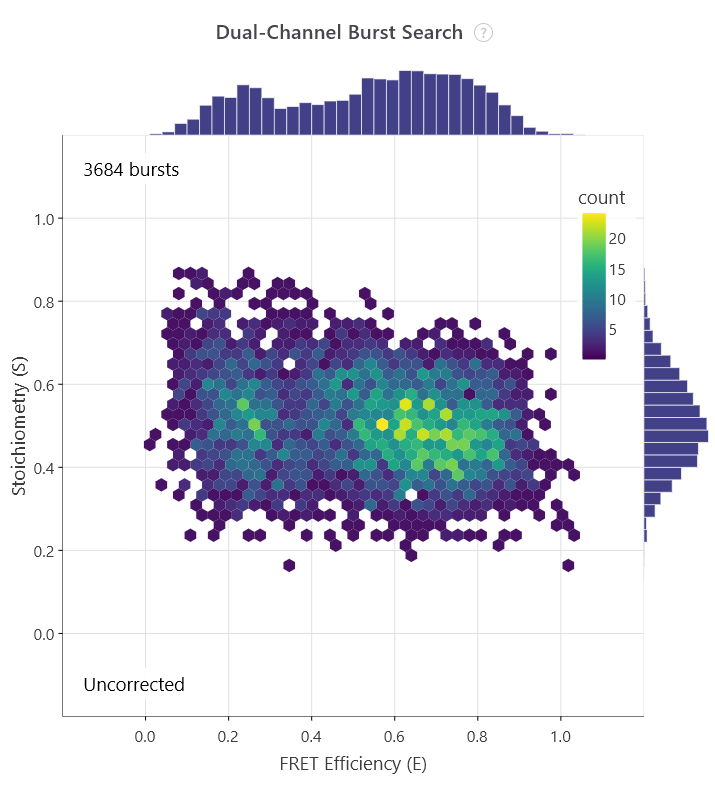
In the same year, the one-dimensional coordinate bottleneck was broken. Hohng, Joo, and Taekjip Ha published ‘Single-molecule three-colour FRET’8. Using a DNA Holliday junction as an experimental system, they engineered a three-dye cascade (Donor, Intermediary, and Acceptor) to monitor two independent distance vectors simultaneously. This structural advancement let biophysicists observe coordinated, non-linear domain movements across complex multi-protein assemblies in real time.
Rewriting the biochemistry textbooks
This expanded biophysical toolkit soon drove major biological discoveries. In 2006, Kapanidis et al. published in Science: ‘Initial transcription by RNA polymerase proceeds through a DNA-scrunching mechanism’9.
For years, the transcription field was deadlocked across three distinct models explaining how RNA polymerase moves during initial transcription: transient excursion, inchworming, or DNA scrunching. By placing site-specific fluorophores on the promoter DNA and the polymerase enzyme, Kapanidis et al. explicitly measured the physical distances during abortive initiation. The data ruled out excursion and inchworming models, visually showing that the static enzyme repeatedly reels in downstream DNA, firmly establishing the ‘DNA scrunching’ mechanism in the textbooks.
Modern specialisation and open science (2010–2020)
With standardised data processing pipelines in place, smFRET entered an exciting era of mechanistic discovery, resolving complex structural questions across the central dogma of life. The technique began tackling multi-subunit, chaotic cellular machinery in native-like environments.
Understanding complex biological mechanisms
This decade proved that smFRET could handle the most notoriously difficult biological systems, setting the stage for industrial scaling:
- In vivo dynamics: Moving past pristine buffers, Sakon and Weninger (2010) achieved a major milestone by tracking single-molecule SNARE conformational transitions upon complex formation inside living cells10
- Complex physiological matrices: Crawford et al. (2013) successfully monitored pre-mRNA structural dynamics during splicing directly within unpurified whole-cell extracts, ensuring these biomolecules were observed in more physiologically relevant environments11
- Transmembrane signalling: Vafabakhsh, Levitz, and Isacoff (2015) used smFRET to probe the activation mechanism of full-length mammalian dimeric G-protein coupled receptors (GPCRs), proving that agonist binding induces a rapid closure of ligand-binding domains followed by a crucial reorientation of the dimer interface12
Unifying analysis standards
Also, by 2010, the field had matured, but was facing highly fractured software analysis pipelines. Laboratories wrote custom, unverified scripts, leading to subtle variance in how different groups calculated background subtraction, cross-talk corrections, and gamma factors.
To transform smFRET into an open, universally reproducible tool, several major standardisation packages were released:
- SMART (2012): Published by Greenfeld et al., the Single Molecule Analysis Research Tool provided an integrated approach for verifying surface-immobilised time trajectories and standardising kinetic state identification using hidden Markov models (HMM)13
- FRETBursts (2016): Developed by Ingargiola et al., FRETBursts established an open-source, robust Python implementation for burst analysis in freely diffusing single-molecule experiments, modernising how background thresholds and burst search algorithms were handled14
- Photon-HDF5 (2016): In another publication from the same year, Ingargiola et al. established an open, standard file format design tailored explicitly for timestamp-based single-molecule architecture. By archiving raw photon data uniformly, it enabled structural data verification across global laboratories15
Democratising access: From open source to benchtop screening (2020–Present)
Despite these extraordinary biological breakthroughs, a steep bottleneck remained for the first 25 years: access.
Historically, conducting an smFRET experiment required custom-built optical instrumentation housed in a dedicated physics darkroom, with an optical table to dampen vibrations, and a specialised team of optics experts just to maintain laser alignment.
The true modern milestone of the smFRET timeline is the dismantling of this barrier.
In 2020, a paradigm shift in instrument accessibility occurred. A collaborative team led by Dr. Timothy Craggs published a pivotal paper in Nature Communications introducing the smfBox – an open-source, benchtop platform for single-molecule FRET16.
The smfBox packaged a highly sensitive, in-solution confocal microscopy workflow into a standardised, radiation-safe, compact box that could operate under ambient room light without an optical table.

While a commercial, custom-built single-molecule instrument traditionally cost upwards of £400,000, the smfBox blueprints allowed academic labs to build their own high-precision instrument for roughly £40,000. Winning the Open Research Prize in 2021, the platform was adopted and constructed by laboratories globally, from the US to South Korea, democratising single-molecule experiments for non-specialists.
The launch of Exciting Instruments and the EI-FLEX Pro
Realising that many laboratories lacked the internal engineering infrastructure to assemble home-builds from blueprints, the foundational team spun out Exciting Instruments in 2021 to fully bridge the gap between academic specialisation and industrial-scale discovery.
The launch of their commercial platforms marked the transition of smFRET from a low-throughput academic specialist technique into a fully integrated, accessible workflow suitable for modern biotechnology and biopharma:
- The EI-FLEX: This platform translated the core ethos of the smfBox into a commercial benchtop solution. Operating entirely under ambient light with intuitive software, it utilises ALEX (Alternating-Laser Excitation) protocols to deliver highly reproducible absolute distance measurements and biomolecular dynamics at sub-nanometre precision

- The EI-FLEX Pro: Representing the cutting edge of high-throughput molecular dynamics, the EI-FLEX Pro builds upon the original EI-FLEX, integrating automated alignment and well-plate compatibility with real-time single-molecule sensitivity

By combining solution-phase confocal smFRET, Fluorescence Correlation Spectroscopy (FCS), and Fluorescence Cross-Correlation Spectroscopy (FCCS) into a unified benchtop platform, these instruments enable the rapid characterisation of complex, dynamic biophysical systems in less than a day.
The launch of the analysis software, PhotonFit, democratises access further. Taking what was an unwieldy series of coding scripts, Exciting Instruments has distilled this complexity into a user-friendly programme that turns raw smFRET and FCS/FCCS data into publication-ready insights with an easy-to-use, real-time workspace designed for all scientists.
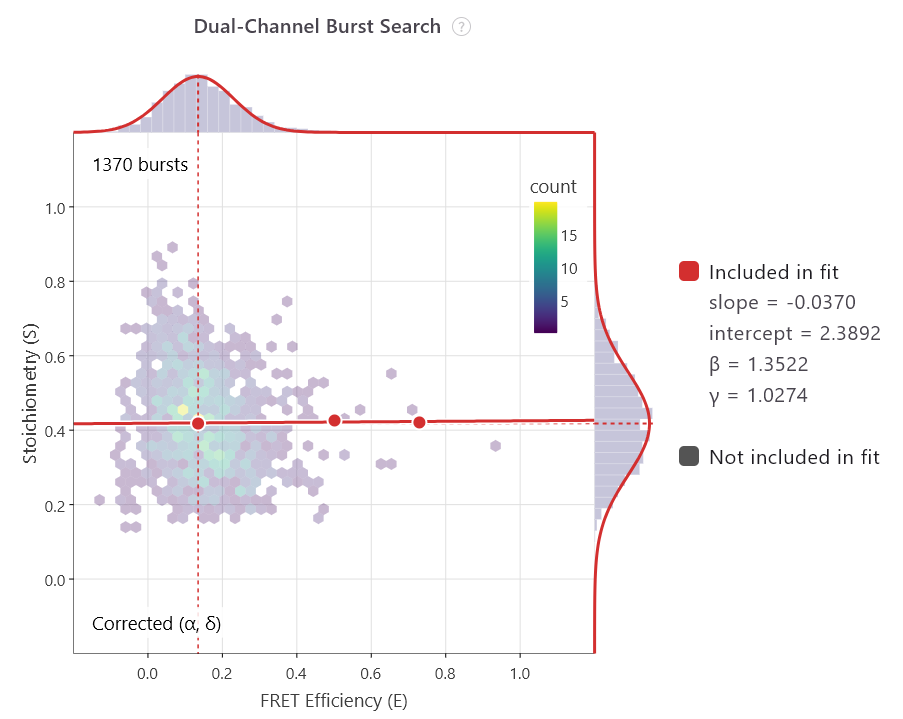
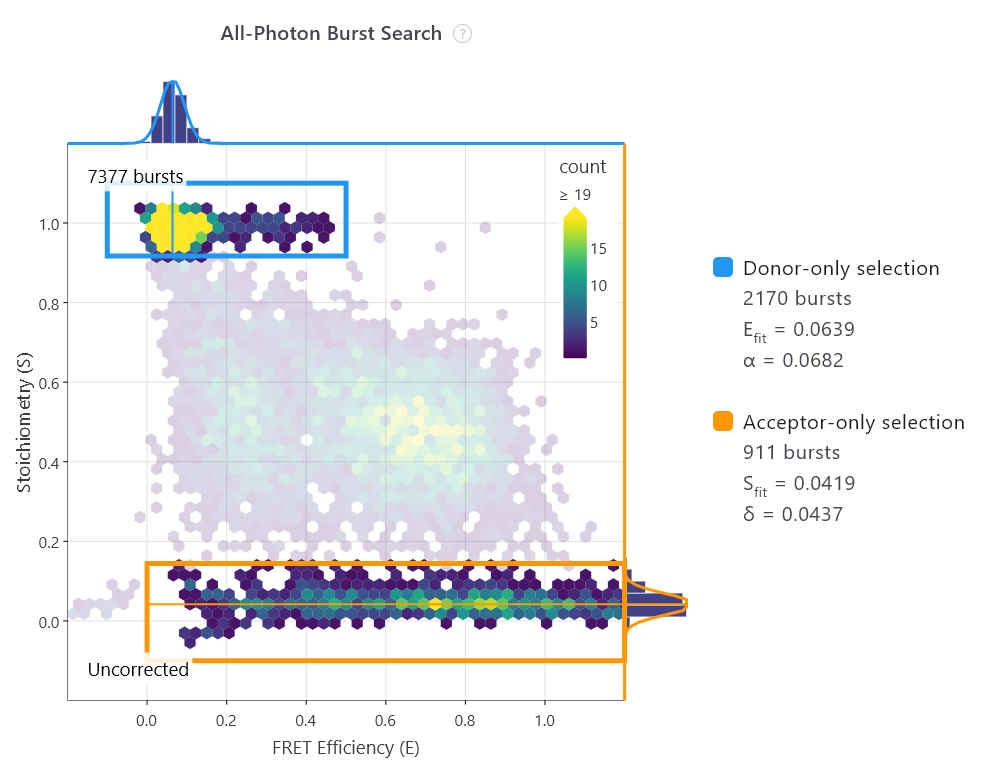


The future of smFRET is exciting
The 30-year trajectory of single-molecule FRET reveals a profound truth about scientific innovation: the ultimate impact of a technology is unlocked only when it is democratised. The first phase of smFRET was defined by establishing single-molecule resolution. The second phase saw the application of that vision to solve biology’s most challenging questions.
As we enter the next 30 years, the focus has fundamentally shifted from ‘can we see a single molecule?’ to ‘what questions can we ask next?’
By packaging decades of biophysical expertise and breakthroughs into an intuitive, benchtop platform, single-molecule biophysics is entering the mainstream. We are moving toward a future where real-time molecular motion is captured in routine workflows, across specialisms and sectors. These will be used to probe the remaining biological questions underpinning life and disease, defining how we engineer the next era of targeted therapeutics and diagnostics.
Simply put, the future of single-molecule FRET is exciting. And since these techniques are no longer confined to the darkrooms of specialised institutes, all you need to get involved is a space on your bench and a biological question.
References
- Stryer, L. & Haugland, R. P. Energy transfer: a spectroscopic ruler. Proceedings of the National Academy of Sciences 58, 719–726 (1967).
- Hirschfeld, T. Optical microscopic observation of single small molecules. Appl. Opt. 15, 2965–2966 (1976).
- Ha, T. et al. Probing the interaction between two single molecules: fluorescence resonance energy transfer between a single donor and a single acceptor. Proceedings of the National Academy of Sciences 93, 6264–6268 (1996).
- Ha, T. et al. Ligand-induced conformational changes observed in single RNA molecules. Proceedings of the National Academy of Sciences 96, 9077–9082 (1999).
- Ha, T. et al. Single-molecule fluorescence spectroscopy of enzyme conformational dynamics and cleavage mechanism. Proceedings of the National Academy of Sciences 96, 893–898 (1999).
- Deniz, A. A. et al. Single-molecule protein folding: Diffusion fluorescence resonance energy transfer studies of the denaturation of chymotrypsin inhibitor 2. Proceedings of the National Academy of Sciences 97, 5179–5184 (2000).
- Kapanidis, A. N. et al. Fluorescence-aided molecule sorting: Analysis of structure and interactions by alternating-laser excitation of single molecules. Proceedings of the National Academy of Sciences 101, 8936–8941 (2004).
- Hohng, S., Joo, C. & Ha, T. Single-Molecule Three-Color FRET. Biophys. J. 87, 1328–1337 (2004).
- Kapanidis, A. N. et al. Initial Transcription by RNA Polymerase Proceeds Through a DNA-Scrunching Mechanism. Science (1979). 314, 1144–1147 (2006).
- Sakon, J. J. & Weninger, K. R. Detecting the conformation of individual proteins in live cells. Nat. Methods 7, 203–205 (2010).
- Crawford, D. J., Hoskins, A. A., Friedman, L. J., Gelles, J. & Moore, M. J. Single-molecule colocalization FRET evidence that spliceosome activation precedes stable approach of 5′ splice site and branch site. Proceedings of the National Academy of Sciences 110, 6783–6788 (2013).
- Vafabakhsh, R., Levitz, J. & Isacoff, E. Y. Conformational dynamics of a class C G-protein-coupled receptor. Nature 524, 497–501 (2015).
- Greenfeld, M., Pavlichin, D. S., Mabuchi, H. & Herschlag, D. Single Molecule Analysis Research Tool (SMART): An Integrated Approach for Analyzing Single Molecule Data. PLoS One 7, e30024- (2012).
- Ingargiola, A., Lerner, E., Chung, S., Weiss, S. & Michalet, X. FRETBursts: An Open Source Toolkit for Analysis of Freely-Diffusing Single-Molecule FRET. PLoS One 11, e0160716- (2016).
- Ingargiola, A., Laurence, T., Boutelle, R., Weiss, S. & Michalet, X. Photon-HDF5: An Open File Format for Timestamp-Based Single-Molecule Fluorescence Experiments. Biophys. J. 110, 26–33 (2016).
- Ambrose, B. et al. The smfBox is an open-source platform for single-molecule FRET. Nat. Commun. 11, 5641 (2020).


