To tether, or not to tether…
The biophysicist’s dilemma
Capturing the fleeting moments of biology presents a fundamental challenge: to observe a molecule, you first have to catch it. A common solution to this problem is to use surface immobilisation. By tethering a biomolecule to a chemically modified glass coverslip, often using biotin-streptavidin or antibody chemistry, researchers can trap a target and observe it over extended periods of time. This approach provides an unparalleled window into long-term kinetics.
But this control comes at a biological cost. Glass coverslips and artificial tethers are dramatically different from the native, three-dimensional fluid environment of a living cell. When we force a molecule onto a solid substrate, we risk introducing significant surface immobilisation artefacts, restricting its natural conformational freedom, or inducing unnatural biases.
This poses a critical dilemma for the modern biophysicist: Do we tether a molecule down to ensure prolonged, controlled observation, or do we let it roam free to ensure biological accuracy?
Surface and solution assays can be viewed as mutually exclusive, although a robust strategy might be to treat them as complementary workflows. In this article we will explore why:
- Including solution-phase assays is vital for eliminating surface artefacts (but comes with its own challenges)
- How techniques like smFRET and FCS are championing biological relevance with biophysical precision
- How combining both modalities provides the definitive, orthogonal data needed to map true-to-life molecular dynamics
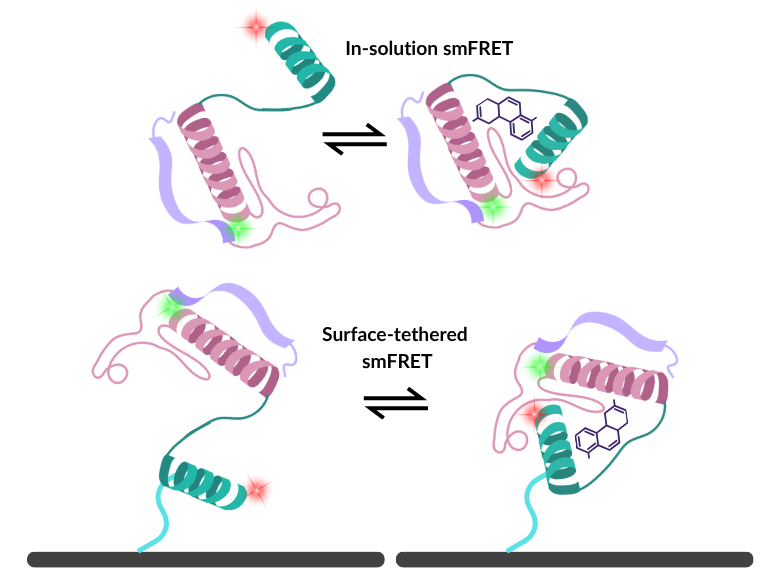
Why tether at all?
For certain experimental designs, anchoring a molecule to a solid substrate is a logistical necessity.
The most profound advantage of surface immobilisation is the extended observation window. When studying slow, complex enzymatic cycles or multi-step conformational transitions that unfold over seconds or minutes, an untethered molecule will diffuse out of a confocal volume too quickly. Techniques like total internal reflection fluorescence (TIRF-FRET) solve this by anchoring target molecules to a glass coverslip, allowing a single biomolecule to be tracked continuously.
Furthermore, a fixed molecule enables complex, multi-step microfluidic manipulation. Because the target is locked in place, researchers can perform wash steps or change ligand concentrations mid-experiment without losing track of the specific molecules under observation. This is one of the advantages of surface plasmon resonance (SPR); many candidate molecules can be tested in parallel by flowing over the same substrate.
The challenges of surface immobilisation
This experimental control comes with a severe caveat: glass coverslips and tethers are a far cry from the crowded, dynamic, three-dimensional fluid environment of a living cell.
When we force a molecule onto a solid substrate, we introduce the risk of significant surface immobilisation artefacts.
Advanced surface passivation techniques (like PEGylation or lipid bilayers) and site-specific tethering chemistries are designed to mitigate these issues, but they do not eliminate them. Even a perfectly site-specifically anchored protein remains in peril; its proximity to the surface can still induce non-specific adsorption, forcing unnatural conformations or denaturing fragile domains.
Even in a best-case scenario where a molecule successfully avoids sticking directly to the substrate, the very act of tethering it introduces persistent steric and rotational restrictions. This surface boundary can mask crucial biological transitions, alter binding kinetics, and physically prevent the molecule from adopting the true, open structures it would naturally exhibit in a free solution. Additionally, while untethered binding partners are freely diffusing in solution, they too can non-specifically adsorb onto the surface, leading to potential false-positive associations.
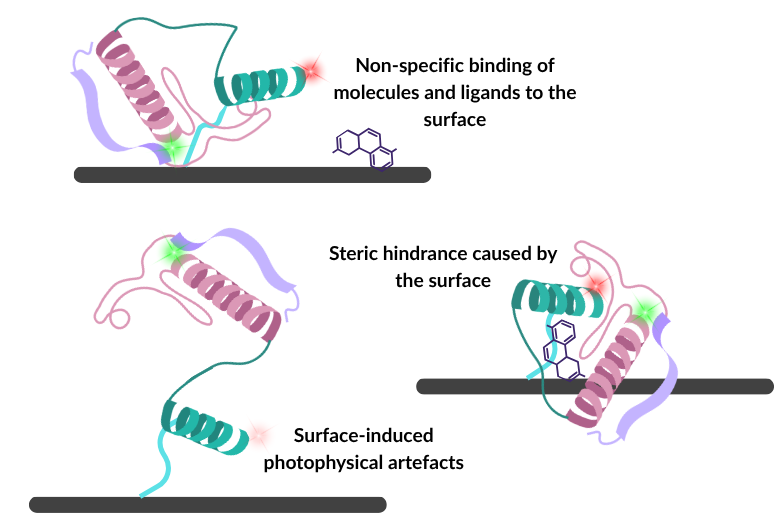
Photophysical artefacts are also potential issues when using immobilised techniques. Tethered labelled biomolecules can cause erratic dye blinking, premature photobleaching, or direct fluorescence quenching. These photophysical anomalies alter the apparent donor-acceptor intensities, leading to artificial shifts in FRET efficiency that do not reflect true conformational states of the biomolecule of interest. Also, given that surface-based approaches are used for prolonged imaging of biomolecules, these undesired photophysical artefacts become more disruptive due to the increased excitation time.
Some molecules simply do not tolerate immobilisation, necessitating a shift in experimental design.
The in-solution alternative
To avoid surface-induced artefacts, many researchers turn to free-diffusion confocal spectroscopy. By focusing a laser into a femtoliter-scale excitation volume, researchers can illuminate labelled biomolecules as they naturally pass through the beam. A precise pinhole blocks out-of-focus light, drastically reducing background fluorescence interference.
Here, molecules retain 100% of their natural rotational and translational freedom. The trade-off is observation time; given that molecules are untethered, they quickly diffuse out of the laser focus, making it impossible to track a single molecule’s long-term history without advanced traps. However, what you lose in longitudinal tracking, you gain in rapid, unrestricted dynamics.
With this in mind, it is important to explore how immobilised techniques are used in research and drug discovery, and examine how orthogonal techniques can support these data.
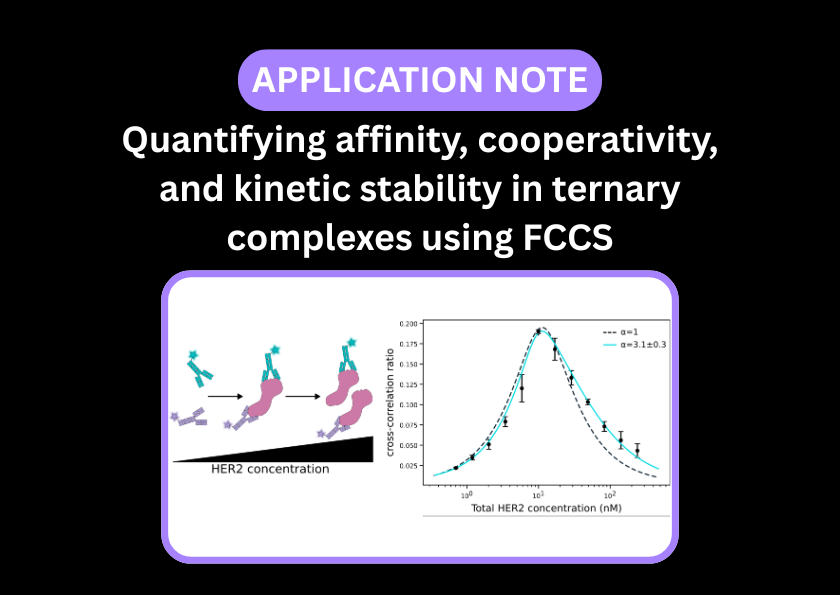
Enhancing the TPD pipeline with solution-phase validation
The excitement around targeted protein degradation (TPD) therapeutics, such as PROTACs and molecular glues, is being reflected in drug discovery pipelines globally. Unlike traditional small-molecule inhibitors that may block an active site, a TPD mechanism involves bringing a target disease-causing protein and an E3 ubiquitin ligase into close proximity. This forms a transient, three-part ternary complex that induces ubiquitination and subsequent degradation of the target.
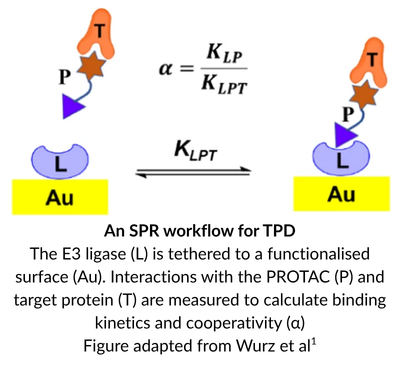
The ultimate goal is to design a small molecule that diverts a target to the protein degradation pathway and reliably results in target destruction. However, mapping ternary complex formation, stability, and kinetics is notoriously difficult. While surface-immobilised biophysical methods like SPR are indispensable for characterising ternary complex candidates, they experience unique spatial challenges when it comes to capturing the native behaviour of these flexible structural assemblies.
In the TPD drug discovery workflow, SPR is used to establish absolute affinities, cooperativity factors, and real-time kinetic rates during candidate screening.
However, to confidently advance a lead compound, a critical distinction must be made between surface-tethered kinetics and true solution-phase behaviour.
When a protein is anchored to an SPR sensor chip, its absolute physical degrees of freedom are fundamentally restricted. For a highly dynamic ternary complex, this surface constraint can be problematic. The physical boundary of the chip surface can obstruct the precise molecular tumbling, loop rearrangements, and three-dimensional search space required for the proteins to explore the range of conformations required to maximise interactions at the binding interface.
While SPR can successfully capture kinetic differences, the physical presence of the surface can artificially blunt or mask absolute cooperativity values and kinetic constants. For advanced lead optimisation, where minor changes to a linker can completely alter the ternary ensemble, validating these parameters in a true, constraint-free solution phase becomes essential to avoid misleading data.
A recent high-impact study from Wurz et al.1 has uncovered structural insights that highlight the dynamic nature of these complexes, exemplifying why ternary complexes require a multi-layered biophysical approach.
They utilised a combination of structural modelling and SPR to demonstrate that a PROTAC’s degradation efficiency is heavily dictated by ternary complex binding affinity and the kinetic half-life of the assembly. The authors used calculated buried surface area (BSA) at the protein-protein interface to structurally validate these experimental parameters.
Maximising this contact zone requires the partner proteins to possess unconstrained spatial mobility so they can dynamically sample different relative orientations. When the E3 ligase is chemically tethered to an SPR chip, its rotation is locked – forcing the incoming target protein to navigate a restricted, single-sided search space to hit the optimal geometry required to maximise BSA.
Direct characterisation for complementary validation
Rather than replacing high-throughput tools like SPR, solution-phase single-molecule techniques should be introduced to research and advanced drug discovery pipelines as highly complementary validators. Researchers can take the top-ranked hits identified by SPR and subject them to deep, unhindered characterisation using two powerful readouts: FCCS and smFRET.
Fluorescence Cross-Correlation Spectroscopy (FCCS) evaluates ternary complex formation under native, unhindered conditions. The target protein is labelled with one colour fluorophore (e.g., green) and the E3 ligase with another (e.g., red). When the PROTAC successfully bridges the two proteins in solution, their co-diffusion can be detected using a two-colour confocal spectrometer, such as the EI-FLEX Pro.
By tracking this co-diffusion, FCCS provides direct observation of unrestricted solution-phase ternary complex formation.
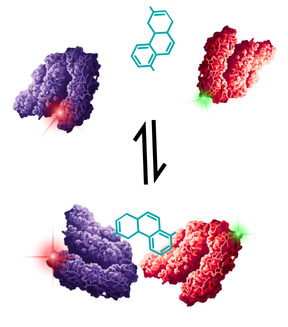
Because the amplitude of the cross-correlation curve directly reflects the concentration of the dual-labelled complex, FCCS allows researchers to cleanly distinguish full ternary complexes from unbound monomers and binary intermediates, free from surface matrix interference.
You can read more on ternary complex characterisation by FCCS in our application note here.
Single-molecule FRET (smFRET) offers highly complementary, structural insights to the binding characterisation of FCCS.
By measuring the distance-dependent energy transfer between two fluorophores, smFRET captures the real-time, solution-phase structural dynamics of the complex. It allows researchers to actively watch how dynamic molecules undergo conformational changes crucial to their function.
Because smFRET operates on free-diffusing molecules, it can reveal whether a PROTAC successfully drives the complex into the highly cooperative, high-BSA conformations predicted by crystallography, or if a compound traps the complex in a non-productive dead-end.
By combining the screening throughput of SPR and the unconstrained solution binding and structural dynamics of FCCS and smFRET, drug discovery teams gain an unmatched, multi-parametric view of their degraders, ensuring that candidate molecules are optimised for the clinic based on physiologically relevant biophysical data.
Why single-molecule resolution is crucial for in-solution methods
Moving away from the surface to utilise in-solution methods does not automatically mean they will be artefact-free. Bulk, in-solution assays like AlphaLISA trade surface artefacts for a different, equally detrimental biochemical phenomenon, known as the Hook Effect – ironically, one of the key parameters that scientists designing PROTACs are trying to characterise.
AlphaLISA relies on bulk proximity energy transfer between donor and acceptor beads. However, if the concentration of your labelled target proteins is too high, they saturate the binding sites on the beads themselves. The beads get crowded out, causing the proximity signal to paradoxically plummet to baseline.
The real nightmare for TPD researchers is that this technological hook effect overlaps directly with the native biological hook effect of the PROTAC (where excess compound saturates proteins independently and prevents ternary complex formation). Trying to untangle whether a dropping signal is caused by the concentration of your therapeutic candidate or your assay beads being saturated needlessly clouds your data.
This is where solution-based assays with single-molecule resolution shine.
FCCS bypasses the requirement for bead-based proximity matrices while measuring the exact ratio of every species in the tube (free monomers, binary intermediates, and full ternary complexes), enabling determination of the optimal dosing window with absolute clarity.
Exploring time and space with complementary single-molecule modalities
The fundamental challenge when mapping the structural dynamics of a biomolecule lies in the fact that biology operates across a massive temporal spectrum. An enzyme might take minutes to complete a multi-step catalytic cycle, but the crucial structural transitions within that cycle happen in microseconds.
Just as in kinetic screening, no single conformational tool can capture the whole picture. Rather than choosing between surface-bound and solution-phase smFRET, the most rigorous structural insights come from treating them as highly complementary, multi-resolution methods.
1. TIRF-smFRET
Surface-immobilised TIRF-smFRET is an exceptional tool for tracking a single molecule’s history over extended periods. Because the molecule is anchored to a glass slide, you can watch it for minutes or hours, making it ideal for mapping the chronological sequence of slow, multi-step pathways.
However, extending the observation window introduces severe trade-offs. Time-lapse imaging is often used to prevent photobleaching of fluorescent dyes, although this drastically lowers temporal resolution and makes it incredibly easy to miss ultra-fast, transient structural states.
Also, to circumvent these photophysical limitations and allow for more continuous imaging, heavy chemical additives like oxygen scavengers and triplet-state quenchers must be introduced. While these cocktails stabilise the dyes, they alter the native chemical microenvironment of the buffer, risking unintended impacts on the stability and behaviour of biomolecules under observation.
2. Confocal smFRET
Fluorescently labelled molecules are observed as they freely diffuse through the confocal volume at thermodynamic equilibrium. smFRET collects thousands of rapid photon bursts, resolving sub-millisecond structural transitions, while also avoiding surface-based artefacts. This technique can capture dynamic conformational changes, alongside Ångstrom-resolution distances between dye positions.
To read more about smFRET and its applications, we recommend our smFRET Handbook.
Can solution-phase techniques capture longer transitions?
Yes, they can – for example, Anti-Brownian Electrokinetic (ABEL) trapping is ideal for bridging the temporal gap between these techniques. It tracks a single diffusing molecule in a microfluidic channel by applying corrective electric fields to counteract its Brownian motion. This holds the molecule inside the confocal laser focus, extending the observation window from milliseconds to several seconds. It gives researchers enough time to track local, unhindered structural dynamics via smFRET without ever introducing a surface tether.
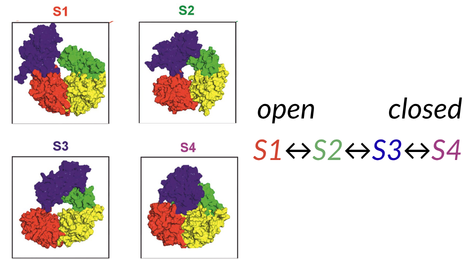
Revealing unknown intermediates – The Rep helicase case study
A premier example of how these techniques can be used together was shown in the study of the bacterial DNA helicase Rep. Howard et al.2 found that their TIRF-FRET data were experiencing photophysical artefacts and so moved to using smFRET to resolve these issues.
By combining the high temporal resolution of free-diffusion confocal smFRET with the extended observation windows of ABEL trapping, the researchers discovered two previously hidden structural intermediates and their interconversion rates.
Conclusion: The power of orthogonal, untethered data
The technological landscape of modern biophysics is broad. No one method will provide all the answers, requiring a multi-angled approach. As therapeutic modalities like targeted protein degradation (TPD) and complex structural systems grow evermore commonplace on our benches, relying on a single experimental format is a distinct scientific risk.
The solution is, well…in-solution.
By treating surface-bound and free-diffusion techniques as complementary methods, you gain the best of both worlds: the massive throughput and chronological tracking of SPR and TIRF, paired with the single-molecule clarity of FCCS and confocal smFRET. Taking your measurements off the surface allows biomolecules to navigate their full spectrum of movement, giving you absolute confidence that your data perfectly mirrors true physiological dynamics.
References
- Wurz, R. P. et al. Affinity and cooperativity modulate ternary complex formation to drive targeted protein degradation. Nat. Commun. 14, 4177 (2023).
- Howard, J. A. L. et al. The transitional kinetics between open and closed Rep structures can be tuned by salt via two intermediate states. Nucleic Acids Res. 54, gkaf1483 (2026).